







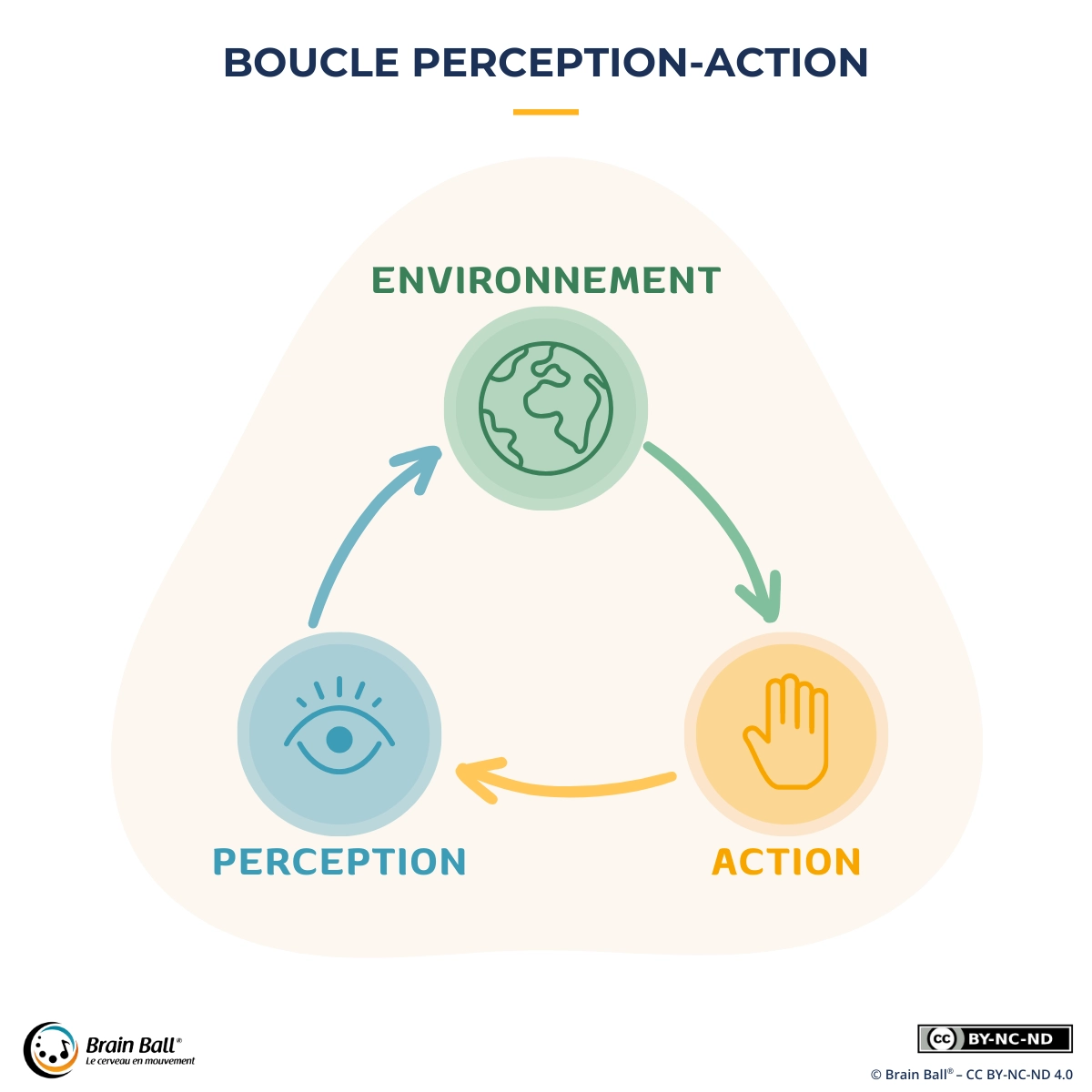
Définition de la boucle perception-action
La boucle perception-action désigne un principe fondamental du contrôle du mouvement et des sciences cognitives : ce que nous percevons guide nos actions, et nos actions transforment en retour ce que nous percevons. Perception et mouvement ne fonctionnent pas en séquence — d’abord percevoir, puis agir — mais en continu, dans un dialogue permanent avec l’environnement et avec notre propre corps. Ce mécanisme en boucle fermée est ce qui rend possibles la coordination, l’adaptation et l’apprentissage moteur en situation réelle, qu’il s’agisse d’attraper une balle, d’éviter un obstacle, d’ajuster son équilibre ou de suivre un rythme.
On désigne ce phénomène également sous le nom de boucle sensori-motrice, terme qui insiste sur l’implication conjointe des systèmes sensoriels — vision, proprioception, toucher, audition, système vestibulaire — et du système moteur. Cette notion est aujourd’hui centrale dans les neurosciences du mouvement, la psychomotricité, les sciences du sport et l’apprentissage moteur.
Gibson, les affordances et la perception orientée vers l’action
Parmi les cadres théoriques qui ont le plus contribué à comprendre la boucle perception-action, la perspective écologique de James J. Gibson est incontournable. Gibson propose une idée en apparence simple mais aux conséquences profondes :
Nous ne percevons pas le monde comme un ensemble abstrait de formes et de couleurs, mais comme un environnement rempli de possibilités d’action.
C’est ici qu’intervient la notion d’affordance (Gibson, 1977, 1979). Une affordance, c’est ce que l’environnement « offre » à un individu en fonction de son corps, de ses capacités et du contexte. Ainsi par exemple, une marche d’escalier offre la possibilité de monter, une poignée celle de tirer, une balle celle d’être saisie ou lancée. Percevoir un objet ou une situation, c’est souvent déjà repérer ce que je peux faire maintenant — la perception est naturellement orientée vers l’action. Cette perspective rompt avec l’idée d’un cerveau qui « traite d’abord » puis « agit ensuite » : chez Gibson, l’organisme et son environnement sont couplés en permanence.
Les approches contemporaines en sciences du sport regroupées sous le terme ecological dynamics (Araújo, Davids & Hristovski, 2006 ; Davids et al., 2013) s’appuient largement sur ce cadre pour expliquer comment les athlètes lisent les affordances en situation et y adaptent leurs décisions motrices en temps réel, sans passer nécessairement par un raisonnement explicite.
Le fonctionnement concrêt de la boucle sensori-motrice : prédiction, action et retour sensoriel
Pour comprendre comment la boucle perception-action opère concrètement, il faut distinguer deux types de mécanismes qui travaillent ensemble : les processus réactifs — le système détecte une erreur et corrige — et les processus prédictifs — le cerveau anticipe les conséquences de ses propres actions avant même d’en recevoir le retour sensoriel.
La description la plus intuitive de la boucle correspond à un contrôle en boucle fermée : l’action se corrige grâce au retour sensoriel. Si la balle arrive plus à gauche que prévu, le bras corrige sa trajectoire ; si l’appui au sol change, la posture se réorganise. Ce retour immédiat — visuel, tactile, proprioceptif — permet une correction continue. C’est cette logique que décrivent notamment Ahissar & Assa (2016), qui proposent de concevoir la perception elle-même comme un processus actif de convergence vers un état cible, plutôt que comme une simple réception d’informations.
Trois exemples pour comprendre la boucle en direct
Lorsque vous attrapez une balle, vous ne faites pas un geste unique. Vous suivez la balle du regard, vous placez la main, puis vous ajustez parce que la balle continue d’avancer et que votre corps bouge aussi. Si la balle arrive légèrement plus à gauche, votre main corrige sa trajectoire. Si elle est plus rapide que prévu, vous adaptez le timing. Au moment du contact, le toucher confirme et vous serrez plus ou moins fort. C’est la boucle perception-action en direct.
Quand vous marchez et évitez un obstacle, vous repérez l’objet au sol, vous modifiez la trajectoire du pied — mais cette modification change votre équilibre. Votre posture se réorganise alors pour rester stable. L’action modifie la perception, et la perception déclenche l’ajustement suivant.
Même écrire mobilise la boucle : l’œil contrôle la forme produite, la main ajuste la pression, le poignet et la posture se réorganisent. Avec l’entraînement, ces corrections deviennent plus automatiques — mais elles restent présentes.
La copie d’efférence : agir avant de percevoir
Mais la boucle ne se réduit pas à des corrections réactives. Lorsque le système nerveux envoie un signal moteur aux muscles, il en adresse simultanément une copie — appelée copie d’efférence — à d’autres structures cérébrales chargées de prédire les conséquences sensorielles attendues de l’action. Cette prédiction permet de comparer ce qui était attendu avec ce qui est réellement perçu, et de détecter des erreurs avec une rapidité que le simple retour sensoriel ne permettrait pas.
Ce mécanisme de prédiction est au cœur de ce que les neurosciences du mouvement appellent les modèles internes — des représentations dynamiques que le cerveau utilise pour simuler les effets de ses propres actions (Wolpert & Kawato, 1998).
Ces modèles se construisent et s’affinent avec l’expérience : plus un mouvement est pratiqué dans des contextes variés, plus les prédictions deviennent précises, et plus les corrections sont rapides et fines.
C’est l’un des mécanismes qui explique pourquoi l’expertise motrice ne ressemble pas à la simple mémorisation d’un geste, mais à une capacité à s’ajuster de façon de plus en plus automatique.
Le rôle du cervelet dans la boucle
Le cervelet est la structure centrale de cette fonction prédictive dans la boucle sensori-motrice. Il compare en permanence les conséquences motrices prédites et les retours sensoriels réels, génère un signal d’erreur, et ajuste la commande motrice en conséquence. Avec la répétition, il contribue à l’automatisation des séquences motrices : les corrections deviennent de moins en moins conscientes, de plus en plus rapides et précises. C’est en partie grâce au cervelet que l’expert peut exécuter des gestes complexes tout en maintenant son attention sur la situation globale — une capacité décisive dans les activités de coordination rythmique ou de jonglage, où la charge attentionnelle est élevée.
Voies visuelles de l’action et neurones miroirs
La vision joue un rôle particulier dans la boucle perception-action, et les neurosciences ont montré qu’elle n’opère pas de façon unitaire. Goodale & Milner (1992) ont proposé une distinction devenue classique entre deux voies visuelles corticales. La voie ventrale (occipito-temporale) traite les propriétés des objets — leur forme, leur identité, leur catégorie — et soutient la perception consciente de l’environnement. La voie dorsale (occipito-pariétale) traite en revanche les informations spatiales et cinématiques nécessaires au guidage de l’action : où est l’objet, comment l’atteindre, dans quelle direction et à quelle vitesse bouger. C’est principalement la voie dorsale qui alimente la boucle perception-action lors des gestes d’atteinte, d’interception ou d’ajustement postural — souvent de façon non consciente et très rapide.
Un autre éclairage sur le lien entre perception et action vient de la découverte des neurones miroirs (Gallese et al., 1996), initialement identifiés chez le macaque dans le cortex prémoteur. Ces neurones s’activent à la fois lors de l’exécution d’une action et lors de l’observation de cette même action réalisée par un congénère. L’hypothèse qui en a découlé — que ce système jouerait un rôle dans la compréhension des actions d’autrui, l’imitation et l’apprentissage social — a suscité un vif intérêt (Rizzolatti & Craighero, 2004). Il convient cependant de préciser que le transfert de ce modèle à l’espèce humaine, et plus encore sa portée fonctionnelle exacte, restent débattus dans la communauté scientifique : Hickok (2009) a formulé des objections substantielles à une interprétation trop large du système miroir chez l’humain, soulignant les limites méthodologiques des données disponibles. L’idée centrale — que perception et action partagent des substrats neuronaux — reste néanmoins un point de convergence solide entre plusieurs traditions de recherche.
Boucle perception-action et cognition incarnée
La boucle perception-action s’inscrit dans une conception plus large du fonctionnement cognitif connue sous le nom de cognition incarnée (embodied cognition). Cette approche, développée notamment par Varela, Thompson & Rosch (1991) puis reprise et nuancée par de nombreux auteurs (Wilson, 2002), soutient que penser, apprendre, mémoriser ou percevoir ne sont pas des processus cérébraux isolés : ils s’appuient sur le corps en action et sur l’interaction continue avec l’environnement.
La boucle perception-action est un point d’entrée concret dans cette idée : elle montre que nos représentations du monde se construisent dans et par le mouvement. Saisir un objet, en estimer le poids, comprendre sa forme, anticiper ce qu’on peut en faire — tout cela passe par des expériences sensorimotrices accumulées. La cognition n’est pas seulement dans la tête ; elle s’enracine dans la façon dont le corps perçoit, se déplace et s’ajuste.
Construction de la boucle : développement et apprentissage moteur
La boucle perception-action ne naît pas constituée : elle se construit et s’affine tout au long du développement. Dès les premières semaines de vie, les nourrissons apprennent à coupler vision et préhension, à calibrer la force de leur saisie, à anticiper les trajectoires d’objets simples. Cette construction repose sur l’expérience motrice active. Une expérience marquante menée par Held et Hein en 1963 le démontre clairement : dix paires de chatons sont élevées dans l’obscurité totale, puis exposées à la lumière uniquement dans un dispositif en forme de carrousel. Dans chaque paire, un chaton peut se déplacer librement et actionne lui-même le mouvement du système. L’autre, placé dans une nacelle, subit exactement le même parcours — même trajectoire, même environnement visuel — mais sans pouvoir contrôler ses déplacements. À la fin de l’expérience, seuls les chatons actifs ont développé une perception visuelle fonctionnelle du mouvement et de l’espace. Les chatons passifs, bien qu’ayant été exposés aux mêmes stimuli visuels, n’y parviennent pas.
En d’autres termes, c’est l’action qui confère à la perception sa dimension fonctionnelle : voir sans agir ne suffit pas à établir le lien entre perception et mouvement.
Chez l’enfant, la manipulation libre d’objets, le jeu moteur et les activités à contraintes variées jouent un rôle essentiel dans ce développement. En psychomotricité, la qualité de la boucle sensori-motrice est souvent un indicateur central de l’intégration du schéma corporel : des difficultés d’ajustement, d’anticipation ou de correction en temps réel peuvent signaler des fragilités dans l’intégration sensorimotrice. Avec l’entraînement et la pratique régulière dans des situations variées, les corrections deviennent progressivement plus automatiques, plus rapides et mieux adaptées — mais elles ne disparaissent jamais : même l’expert corrige en permanence.
La boucle perception-action dans le sport et la performance
Dans le sport, l’environnement change en permanence : les adversaires et partenaires se déplacent, les trajectoires sont imprévisibles, le temps d’action est limité. La performance dépend alors fortement de la capacité à coupler rapidement et efficacement perception et action.
Une boucle perception-action bien entraînée permet d’abord de gagner du temps : l’athlète expert repère plus tôt les indices pertinents, anticipe plus tôt, ajuste sans délai. Elle permet aussi de réussir malgré la variabilité — un rebond inattendu, une passe imparfaite, un changement de rythme. Elle contribue enfin à la régularité : l’expert n’est pas celui qui ne se trompe jamais, mais celui qui corrige mieux et plus vite. Les approches en ecological dynamics (Araújo et al., 2006 ; Davids et al., 2013) insistent précisément sur ce couplage et sur la lecture des affordances en situation comme fondements de l’efficacité motrice en contexte réel.
La variabilité des conditions d’entraînement joue ici un rôle central. En variant les paramètres — vitesse, distance, trajectoire, contraintes temporelles — on entraîne le système à comprendre la logique du geste plutôt qu’à copier une forme figée. Le résultat est une capacité d’adaptation transférable à des situations nouvelles, y compris hors du contexte sportif.
Le rythme comme régulateur de la boucle : synchronisation sensorimotrice
Une dimension moins souvent abordée dans les descriptions classiques de la boucle perception-action est son articulation avec le rythme. Lorsqu’un individu synchronise ses mouvements avec un stimulus externe — un métronome, un tempo musical — la boucle sensori-motrice prend une forme particulièrement précise : le système nerveux doit en permanence comparer la position temporelle de ses gestes avec le signal de référence, détecter les décalages, et les corriger.
Les recherches de Repp (2005) et Repp & Su (2013) sur la synchronisation sensorimotrice ont mis en évidence deux types d’ajustements que l’organisme effectue plus ou moins consciemment. La correction de phase consiste à recaler le geste sur le temps fort le plus proche lorsqu’un décalage ponctuel s’est produit. La correction de période consiste à recalibrer progressivement le tempo interne quand une dérive s’installe au fil du temps. Ces deux mécanismes montrent que la synchronisation n’est pas un état passif et figé, mais un processus actif et continu de régulation — une boucle dans la boucle.
Ce couplage entre rythme externe et mouvement mobilise des réseaux auditivo-moteurs dont l’importance a été documentée dans plusieurs travaux récents. Patel & Iversen (2014), avec le modèle ASAP (Action Simulation for Auditory Prediction), proposent que la perception d’un rythme repose sur un couplage entre les aires auditives et celles dédiées à la planification motrice — principe au cœur de la stimulation rythmique auditive. Le rythme agit ainsi comme un cadre temporel structurant pour la boucle perception-action : il organise les fenêtres d’anticipation, réduit l’incertitude temporelle et améliore la précision des corrections. C’est en partie ce mécanisme qui explique pourquoi les activités rythmiques — marche cadencée, danse, jonglage en musique — améliorent la qualité du contrôle sensorimoteur, y compris chez des personnes présentant des difficultés d’intégration.
Pour approfondir la question du rythme et de son action sur le cerveau en mouvement, voir notre article sur Robert Zatorre, dont les travaux sur le couplage auditivo-moteur éclairent directement ces mécanismes.
❯
Pour aller plus loin, lire notre article : Zatorre : la musique, le cerveau et le plaisir de bouger
Ce que le Brain Ball® apporte à la boucle perception-action
Le Brain Ball® est conçu pour solliciter la boucle perception-action de façon multisensorielle, rythmique et progressive. Plusieurs mécanismes sont à l’œuvre simultanément, et leur combinaison crée des conditions d’apprentissage particulièrement favorables au développement du contrôle sensorimoteur.
La balle en rebond fournit un feedback immédiat : trajectoire, erreur, réussite — tout est visible et ressenti instantanément. La pratique contraint le participant à corriger en temps réel, ce qui entraîne précisément les mécanismes de détection d’erreur et d’ajustement décrits dans les sections précédentes. Le rythme imposé par le tempo musical ou le métronome ajoute une contrainte temporelle : il ne suffit pas de réussir le geste, il faut le réussir au bon moment. Ce sont les mécanismes de correction de phase et de période qui sont ainsi sollicités — l’animateur structure une synchronisation sensorimotrice active et répétée.
La richesse sensorielle de la pratique renforce encore la boucle : la vision suit la trajectoire de la balle, la proprioception stabilise la posture et informe sur la position des membres, le toucher confirme le contact avec l’objet, et l’audition du tempo cale l’action dans le temps. Ces modalités ne fonctionnent pas en parallèle mais en interaction : chacune contribue à affiner les prédictions et les corrections du système nerveux. En ce sens, Brain Ball® exploite pleinement la logique des modèles internes décrits par Wolpert & Kawato (1998) : chaque séance enrichit les représentations prédictives du mouvement.
La dimension collective de certains exercices ajoute une couche supplémentaire : observer un partenaire, anticiper son geste, s’accorder avec lui mobilise les circuits d’action-observation évoqués plus haut, et fait de la boucle perception-action un phénomène social autant que moteur.
Enfin, la variabilité des exercices — changements de rythme, de matériel, de configuration — est un levier d’apprentissage central : en variant les conditions, Brain Ball® empêche la simple mémorisation d’un geste figé et entraîne une capacité d’adaptation transférable à d’autres contextes de vie.
À retenir
La boucle perception-action décrit le dialogue continu entre ce que nous percevons et ce que nous faisons : percevoir pour agir, et agir pour mieux percevoir. Cette boucle implique à la fois des mécanismes réactifs (correction à partir du retour sensoriel) et des mécanismes prédictifs (copie d’efférence, modèles internes), avec le cervelet comme organe central de la comparaison entre prédiction et réalité. Les affordances de Gibson (1977) expliquent pourquoi la perception est naturellement orientée vers l’action. Les voies visuelles dorsale et ventrale (Goodale & Milner, 1992) montrent comment le cerveau traite séparément la reconnaissance des objets et le guidage du mouvement. La cognition incarnée (Varela et al., 1991) rappelle que nos capacités cognitives se construisent largement dans et par le mouvement. Dans le domaine rythmique, les mécanismes de correction de phase et de période (Repp, 2005 ; Repp & Su, 2013) montrent que la synchronisation avec un stimulus externe est elle-même une forme active de boucle sensori-motrice. Le Brain Ball® exploite l’ensemble de ces mécanismes : feedback immédiat, contrainte rythmique, richesse sensorielle et variabilité des situations.
Pour aller plus loin :
Ahissar, E., & Assa, E. (2016). Perception as a closed-loop convergence process. eLife, 5, e12830. https://doi.org/10.7554/eLife.12830
Araújo, D., Davids, K., & Hristovski, R. (2006). The ecological dynamics of decision making in sport. Psychology of Sport and Exercise, 7(6), 653–676. https://doi.org/10.1016/j.psychsport.2006.07.009
Davids, K., Araújo, D., Vilar, L., Renshaw, I., & Pinder, R. (2013). An ecological dynamics approach to skill acquisition: Implications for development of talent in sport. Sports Medicine, 43(3), 157–167. https://doi.org/10.1007/s40279-012-0001-z
Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Action recognition in the premotor cortex. Brain, 119(2), 593–609. https://doi.org/10.1093/brain/119.2.593
Gibson, J. J. (1977). The theory of affordances. In R. Shaw & J. Bransford (Eds.), Perceiving, acting, and knowing (pp. 67–82). Lawrence Erlbaum Associates.
Gibson, J. J. (1979). The ecological approach to visual perception. Houghton Mifflin.
Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways for perception and action. Trends in Neurosciences, 15(1), 20–25. https://doi.org/10.1016/0166-2236(92)90344-8
Held, R., & Hein, A. (1963). Movement-produced stimulation in the development of visually guided behavior. Journal of Comparative and Physiological Psychology, 56(5), 872–876. https://doi.org/10.1037/h0040546
Hickok, G. (2009). Eight problems for the mirror neuron theory of action understanding in monkeys and humans. Journal of Cognitive Neuroscience, 21(7), 1229–1243. https://doi.org/10.1162/jocn.2009.21189
Patel, A. D., & Iversen, J. R. (2014). The evolutionary neuroscience of musical beat perception: The Action Simulation for Auditory Prediction (ASAP) hypothesis. Frontiers in Systems Neuroscience, 8, 57. https://doi.org/10.3389/fnsys.2014.00057
Repp, B. H. (2005). Sensorimotor synchronization: A review of the tapping literature. Psychonomic Bulletin & Review, 12(6), 969–992. https://doi.org/10.3758/BF03206433
Repp, B. H., & Su, Y.-H. (2013). Sensorimotor synchronization: A review of recent research (2006–2012). Psychonomic Bulletin & Review, 20(3), 403–452. https://doi.org/10.3758/s13423-012-0371-2
Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system. Annual Review of Neuroscience, 27, 169–192. https://doi.org/10.1146/annurev.neuro.27.070203.144230
Varela, F. J., Thompson, E., & Rosch, E. (1991). The embodied mind: Cognitive science and human experience. MIT Press.
Wilson, M. (2002). Six views of embodied cognition. Psychonomic Bulletin & Review, 9(4), 625–636. https://doi.org/10.3758/BF03196322
Wolpert, D. M., & Kawato, M. (1998). Multiple paired forward and inverse models for motor control. Neural Networks, 11(7–8), 1317–1329. https://doi.org/10.1016/S0893-6080(98)00066-5
FAQ sur la Boucle perception-action
Comment la boucle perception-action se différencie-t-elle du simple réflexe ?
Peut-on entraîner la boucle perception-action ?
Pourquoi la variabilité aide-t-elle à apprendre ?
Parce qu’elle pousse le cerveau et le corps à comprendre la logique du geste, pas à copier un mouvement. Quand les conditions changent (vitesse, distance, rythme, trajectoire, consigne, partenaire), on est obligé de repérer les indices vraiment utiles et de s’ajuster : dosage de la force, timing, posture, trajectoire, regard.
Résultat : on n’apprend pas seulement “un mouvement”, on apprend une capacité d’adaptation. Et c’est précisément cette adaptabilité qui rend l’apprentissage plus solide et plus transférable : on réussit non seulement dans l’exercice, mais aussi quand la situation réelle varie (sport, classe, vie quotidienne).
La boucle perception-action est-elle universelle chez les espèces ?
