Définition de la boucle perception-action
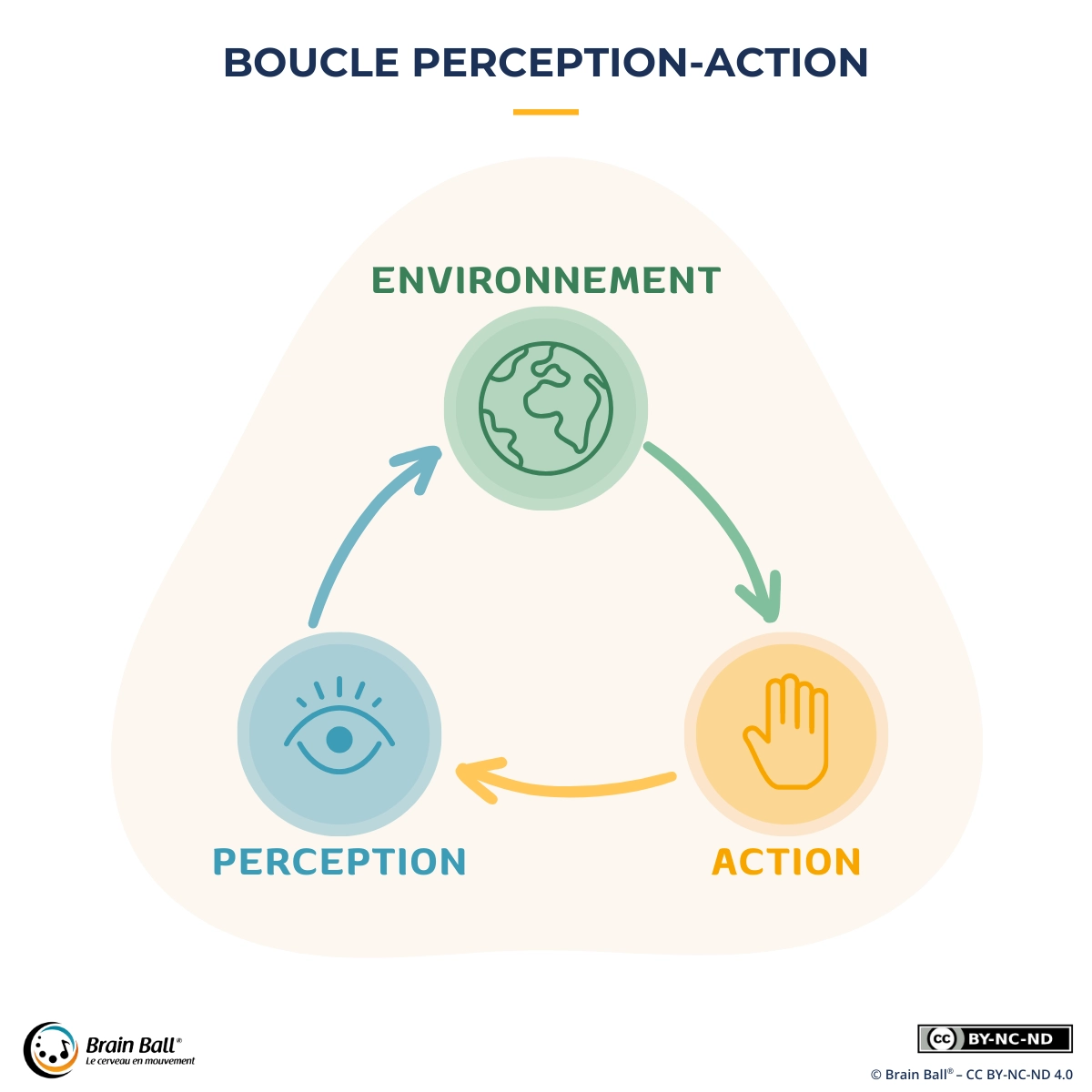
La boucle perception-action désigne une idée simple mais essentielle : ce que nous percevons guide nos actions, et nos actions transforment immédiatement ce que nous percevons. Dans la vie quotidienne, perception et mouvement ne fonctionnent donc pas “à la suite” l’un de l’autre (je perçois puis j’agis), mais ensemble, dans un va-et-vient continu.
Chaque fois que vous bougez votre regard, votre tête, votre main ou votre posture, vous modifiez les informations disponibles. Et dès que ces informations changent (distance, trajectoire, équilibre, rythme, intention d’un partenaire), votre action s’ajuste. C’est ce mécanisme en boucle qui rend possibles la coordination, l’apprentissage et l’adaptation en situation réelle.
Origines et fondements conceptuels : Gibson, affordances et neurones miroirs
Parmi les apports majeurs pour comprendre la boucle perception-action, la perspective écologique de James J. Gibson occupe une place centrale. Gibson propose une idée très intuitive pour le grand public : nous ne percevons pas le monde uniquement comme un décor fait de formes et de couleurs, mais comme un environnement rempli de possibilités d’action.
C’est ici qu’intervient la notion d’affordance (Gibson, 1977). Une affordance, c’est ce que l’environnement “offre” à un individu en fonction de son corps et du contexte. Une marche “offre” la possibilité de monter, une poignée celle de tirer, une balle celle d’être attrapée ou lancée. Autrement dit, la perception est naturellement orientée vers l’action : dans de nombreuses situations, « voir » revient déjà à repérer “ce que je peux faire maintenant”, et « agir » revient à explorer et affiner ce que je perçois.
Un autre exemple marquant pour illustrer le lien entre observation et action est celui des neurones miroirs (Gallese et al., 1996). L’idée centrale ? Certains circuits cérébraux, normalement sollicités lors de l’exécution d’une action, s’activent également — bien que de manière atténuée — lorsque nous observons cette même action chez autrui.
Ce mécanisme jouerait un rôle clé dans notre capacité à comprendre les intentions d’autrui, à anticiper ses mouvements, et même à apprendre par imitation (Rizzolatti & Craighero, 2004). Une fois de plus, cette découverte confirme que perception et action ne sont pas des processus isolés : elles s’influencent mutuellement, en temps réel.
Que se passe-t-il concrètement dans une boucle sensori-motrice ?
Pour se représenter la boucle perception-action, il est utile de penser en termes de boucle sensori-motrice : un dialogue permanent entre ce que les sens captent et ce que le corps fait. Tout commence par une information pertinente, par exemple une trajectoire, une distance, une position de main, un rythme, un appui au sol. Le cerveau initie alors une action : déplacer le regard, tendre le bras, réorienter le bassin, ajuster la force, accélérer ou freiner.
À peine l’action commence-t-elle qu’elle produit un retour immédiat : vous voyez si la balle dévie, vous sentez un déséquilibre, vous percevez un décalage de timing, vous ressentez le contact. Ce retour permet une correction rapide : une micro-rotation du poignet, un pas d’ajustement, une ouverture de main un peu plus tôt, un dosage de force plus fin. La boucle tourne ainsi en continu, souvent très vite, et c’est précisément cette capacité à corriger en temps réel qui fait la différence entre “faire un geste” et “le réussir en situation”.
Cette logique “en boucle fermée” est d’ailleurs un point important dans certains travaux qui décrivent la perception comme un processus actif d’ajustement continu plutôt que comme une simple réception d’informations (Ahissar & Assa, 2016).
Exemples simples pour comprendre la boucle
Lorsque vous attrapez une balle, vous ne faites pas un geste unique. Vous suivez la balle du regard, vous placez la main, puis vous ajustez parce que la balle continue d’avancer et que votre corps bouge aussi. Si la balle arrive légèrement plus à gauche, votre main corrige sa trajectoire. Si elle est plus rapide que prévu, vous adaptez le timing. Au moment du contact, le toucher confirme et vous serrez plus ou moins fort. C’est la boucle perception-action “en direct”.
Quand vous marchez et évitez un obstacle, vous repérez l’objet au sol, vous modifiez la trajectoire du pied, mais cette modification change votre équilibre. Votre posture se réorganise alors pour rester stable. L’action modifie la perception, et la perception déclenche l’ajustement suivant.
Même écrire mobilise la boucle : l’œil contrôle la forme produite, la main ajuste la pression, le poignet et la posture se réorganisent. Avec l’entraînement, ces corrections deviennent plus automatiques, mais elles restent présentes.
Lien entre boucle perception-action et cognition incarnée
La boucle perception-action rejoint directement une idée de plus en plus présente dans les sciences cognitives : la cognition incarnée (ou “embodied cognition”). Cette approche rappelle que penser, apprendre, décider ou se concentrer ne se réduit pas à “faire tourner” un cerveau isolé ; ces processus s’appuient aussi sur le corps en action et sur l’interaction avec l’environnement (Varela et al., 1991 ; Wilson, 2002).
Dit simplement : une partie de nos capacités cognitives se construit et se manifeste dans la manière dont nous bougeons, percevons, manipulons, anticipons et nous ajustons. La boucle perception-action est donc un excellent point d’entrée pour comprendre la cognition incarnée, parce qu’elle montre concrètement comment perception, mouvement et ajustements participent à la compréhension de la situation et à l’efficacité de l’action.
L’intérêt de la boucle perception-action dans le sport
Dans le sport, l’environnement est rarement stable : adversaires, partenaires, vitesse, fatigue, pression, trajectoires parfois imprévisibles. La performance dépend donc fortement de la capacité à coupler perception et action.
Une boucle perception-action efficace aide d’abord à gagner du temps : l’athlète repère plus vite les indices utiles, anticipe plus tôt, ajuste sans retard. Elle permet aussi de réussir malgré la variabilité : une passe imparfaite, un rebond inattendu, un changement de rythme, une feinte. Enfin, elle contribue à la régularité : l’expert n’est pas celui qui ne se trompe jamais, mais celui qui corrige mieux, plus vite, et stabilise progressivement ses solutions. Les approches “écologiques” et les modèles d’“écological dynamics” en sport insistent justement sur ce couplage perception-action et sur la lecture des affordances en situation (Araújo et al., 2006 ; Davids et al., 2013).
Lien et apport du Brain Ball
Le Brain Ball® s’inscrit naturellement dans la logique de la boucle perception-action, parce que la pratique met les participants dans un contexte où ils doivent percevoir, agir, recevoir un retour et ajuster, avec des objets en mouvement et un rythme structurant.
La balle crée un feedback immédiat : trajectoire, erreur, réussite, tout est visible et ressenti tout de suite. Le rythme ajoute une contrainte temporelle : il ne suffit pas de réussir, il faut réussir au bon moment, ce qui renforce les ajustements de timing et de coordination. La richesse multisensorielle renforce encore la boucle : la vision guide, la proprioception stabilise, le toucher confirme, l’écoute du tempo cale l’action, et le corps s’organise comme un ensemble. Enfin, la dimension collective (observer un partenaire, s’accorder, s’adapter à l’autre) alimente en continu le lien entre perception et action, tout en rendant l’activité plus motivante et engageante.
En pratique, le Brain Ball rend la boucle perception-action très concrète : on voit ce qui se passe, on ressent l’ajustement nécessaire, on corrige, on stabilise… et on progresse parce que le corps apprend à s’adapter dans des situations variées, proches du réel.
À retenir
La boucle perception-action explique pourquoi perception et mouvement sont indissociables : on perçoit pour agir, et on agit pour mieux percevoir. Les notions d’affordance (Gibson, 1977) et de neurones miroirs (Gallese et al., 1996) aident à comprendre comment l’environnement et l’observation soutiennent l’action et l’apprentissage. Cette boucle est particulièrement décisive dans le sport, où réussir signifie souvent s’adapter plus que répéter. Elle fait aussi le pont avec la cognition incarnée, en montrant que nos capacités cognitives se construisent largement dans l’action. Le Brain Ball s’appuie précisément sur ces principes en sollicitant en continu : perception, ajustements, rythme, feedback et coordination.
Pour aller plus loin :
- Ahissar, E., & Assa, E. (2016). Perception as a closed-loop convergence process. eLife, 5, e12830. https://doi.org/10.7554/eLife.128
- Araújo, D., Davids, K., & Hristovski, R. (2006). The ecological dynamics of decision making in sport. Psychology of Sport and Exercise, 7(6), 653–676. https://doi.org/10.1016/j.psychsport.2006.07.009
- Davids, K., Araújo, D., Vilar, L., Renshaw, I., & Pinder, R. (2013). An ecological dynamics approach to skill acquisition: Implications for development of talent in sport. Sports Medicine, 43(3), 157–167.
- Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Action recognition in the premotor cortex. Brain, 119(2), 593–609. https://doi.org/10.1093/brain/119.2.593
- Gibson, J. J. (1977). The theory of affordances. In R. Shaw & J. Bransford (Eds.), Perceiving, acting, and knowing (pp. 67–82). Lawrence Erlbaum Associates.
- Gibson, J. J. (1979). The ecological approach to visual perception. Houghton Mifflin.
- Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system. Annual Review of Neuroscience, 27, 169–192. https://doi.org/10.1146/annurev.neuro.27.070203.144230
- Varela, F. J., Thompson, E., & Rosch, E. (1991). The embodied mind: Cognitive science and human experience. MIT Press.
- Wilson, M. (2002). Six views of embodied cognition. Psychonomic Bulletin & Review, 9(4), 625–636. https://doi.org/10.3758/BF03196322
FAQ sur la Boucle perception-action
Comment la boucle perception-action se différencie-t-elle du simple réflexe ?
Peut-on entraîner la boucle perception-action ?
Pourquoi la variabilité aide-t-elle à apprendre ?
Parce qu’elle pousse le cerveau et le corps à comprendre la logique du geste, pas à copier un mouvement. Quand les conditions changent (vitesse, distance, rythme, trajectoire, consigne, partenaire), on est obligé de repérer les indices vraiment utiles et de s’ajuster : dosage de la force, timing, posture, trajectoire, regard.
Résultat : on n’apprend pas seulement “un mouvement”, on apprend une capacité d’adaptation. Et c’est précisément cette adaptabilité qui rend l’apprentissage plus solide et plus transférable : on réussit non seulement dans l’exercice, mais aussi quand la situation réelle varie (sport, classe, vie quotidienne).
La boucle perception-action est-elle universelle chez les espèces ?
